Модификации генетических алгоритмов
3.7. Генетические микроалгоритмы
Данная модификация предназначена для решения задач, которые не требуют популяций с большим числом особей и длинных хромосом. Такой подход оправдан в том случае, когда решение (не обязательно глобальный оптимум) необходимо найти как можно быстрее. В этом случае необходимо уменьшить трудоемкость, связанную с большим количеством итераций, ценой возможно ухудшения качества решения. Такой подход, например, реализован в программе FlexTool [4] следующим генетическим микроалгоритмом.
- Формирование популяции из 5 особей. При этом можно либо случайным образом выбирать все 5 хромосом, либо сохранять одну "хорошую" особь, полученную на предыдущих итерациях, и случайным образом генерировать остальные 4 особи.
- Расчет значений фитнесс-функции особей в популяции и выбор лучшей особи. Присвоить ей номер 5 и перенести в следующее поколение (согласно элитарной стратегии).
- Выбор для репродукции остальных четырех хромосом на основе турнирного метода селекции. При этом хромосомы группируются случайным образом, и соседние пары соперничают за оставшиеся 4 места. Необходимо следить, чтобы родительская пара не формировалась из двух копий одной и той же хромосомы.
- Выполнение кроссинговера с вероятностью
 (вероятность мутации положить
(вероятность мутации положить  ).
). - Проверка сходимости алгоритма (на основе сравнения фенотипов или генотипов). Если условие останова не выполнено, то переход на шаг 2, иначе конец.
Отметим, что в генетическом микроалгоритме используется небольшой фиксированный размер популяции и элитарная стратегия отбора родителей, которая предотвращает потерю хороших особей. Мутация здесь не применяется поскольку разнообразие генетического материала обеспечивается формированием новой популяции при каждом рестарте алгоритма( переходе на шаг 1 при обнаружении сходимости). Процедуры "старта" и "рестарта" предназначены для предотвращения преждевременной сходимости.
3.8. Генетические алгоритмы с изменяемой мощностью популяции
Мощность популяции 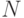 является важнейшим параметром ГА, который критичен во многих приложениях. Если мало, то ГА работает быстро, но при этом увеличивается опасность преждевременной сходимости к локальному экстремуму. Большая мощность популяции увеличивает генофонд, но процесс поиска замедляется.
является важнейшим параметром ГА, который критичен во многих приложениях. Если мало, то ГА работает быстро, но при этом увеличивается опасность преждевременной сходимости к локальному экстремуму. Большая мощность популяции увеличивает генофонд, но процесс поиска замедляется.
На разных этапах работы ГА оптимальное значение может быть различным. На начальном этапе должно быть большим, а на заключительном можно уменьшить.
При одном из подходов в ГА с изменяемым размером популяции каждой особи после ее рождения на текущем этапе оценки целевой функции (ЦФ) присваивается "время жизни" (life time)  – параметр, зависящий от ЦФ особи. Таким образом, каждая особь живет определенное число поколений и умирает по окончании срока жизни. Очевидно, значение этого параметра влияет на размер популяции. В этом случае ГА можно реализовать, например, следующим образом [5].
– параметр, зависящий от ЦФ особи. Таким образом, каждая особь живет определенное число поколений и умирает по окончании срока жизни. Очевидно, значение этого параметра влияет на размер популяции. В этом случае ГА можно реализовать, например, следующим образом [5].
Нестационарный_ГА
{
t =0;
Инициализация;
Оценка ЦФ p(t);
While (условие окончания не выполнено)
{
t=t+1;
Увеличение возраста каждой особи на 1;
Рекомбинация p(t):
Мутация p(t):
Оценка ЦФ p(t):
Определение срока жизни особей;
Удаление из p(t) всех особей с возрастом больше срока жизни;
}
}
Здесь в текущем поколении 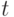 алгоритм обрабатывает популяцию
алгоритм обрабатывает популяцию  . В процессе рекомбинации и мутации генерируется промежуточная популяция, состоящая из потомков, и ее размер пропорционален числу исходной
. В процессе рекомбинации и мутации генерируется промежуточная популяция, состоящая из потомков, и ее размер пропорционален числу исходной 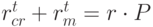 .
.
Тогда 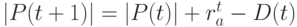 – число особей в новой популяции.
– число особей в новой популяции.
Срок жизни для каждой особи определяется после оценки ЦФ (по формулам, приведенным ниже) и является окончательным, то есть постоянным в процессе эволюции. В таком случае особь живет определенное число поколений, а затем умирает. Срок жизни определяет число поколений, в течении которых особь держится в популяции. Очевидно, что в этом случае, чем больше срок жизни, тем больше потомков может дать особь, так как на каждом этапе родители для рекомбинации выбираются случайно с равной вероятностью. При этом подходе важнейшую роль играет метод определения срока жизни особи.
Очевидно, постоянное значение 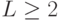 для каждой особи ведет к экспоненциальному росту размеров популяции. Поэтому для каждой особи срок жизни вычисляется индивидуально в зависимости от значения ее ЦФ.
для каждой особи ведет к экспоненциальному росту размеров популяции. Поэтому для каждой особи срок жизни вычисляется индивидуально в зависимости от значения ее ЦФ.
Наиболее часто используются следующие три способа определения срока жизни.
Для их определения введем следующие обозначения:
-
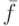 - среднее значение ЦФ по популяции;
- среднее значение ЦФ по популяции; -
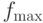 - максимальное значение ЦФ по популяции;
- максимальное значение ЦФ по популяции; -
 - минимальное значение ЦФ по популяции;
- минимальное значение ЦФ по популяции; -
 - абсолютное максимальное значение;
- абсолютное максимальное значение; -
 - абсолютное минимальное значение;
- абсолютное минимальное значение; -
 - максимальный срок жизни;
- максимальный срок жизни; -
 - минимальный срок жизни.
- минимальный срок жизни.
- При пропорциональном методе определения срока жизни:
![L_f=\min\left\{L_{min} +\eta\cdot\frac{f[i]}{\bar f},L_{max}\right\}](/sites/default/files/tex_cache/1dbf2734af2b164f48e36ef398af6e5c.png)
( 3.13) где
 (используется для всех формул).
(используется для всех формул). - При линейном методе определения срока жизни
![L_f=L_{min}+2\cdot\eta\cdot\frac{f[i]-f_{a\min}}{f_{a\max}-f_{a\min}}](/sites/default/files/tex_cache/6280923f271c7e185ea2df16cca2c7a8.png)
( 3.14) - В билинейном методе определения срока жизни
![L_f=\begin{cases}L_{\min} +\eta \cdot \frac {f[i]-f_\min }{\bar f-f_\min },&\text{если $\bar f\ge f[i]$}\\\frac {1}{2}\cdot (L_{\min} +L_{\max} )+\eta \cdot \frac {f[i]-\bar f}{f_\max - \bar f},&\text{если $\bar f< f[i]$}\end{cases}](/sites/default/files/tex_cache/50e6bfae682bea49ca0c371016633711.png)
( 3.15)
Очевидно, что первый метод соответствует пропорциональному отбору отбора родителей - "рулетке". К минимальному сроку жизни добавляется премиальный срок, который пропорционален значению ЦФ для данной особи. Однако эта стратегия имеет серьезный недостаток – она не учитывает информацию о некоторых объективных характеристиках особи, такой, например, как отношение ![\frac{f[i]}{f_\max}](/sites/default/files/tex_cache/c89b6022d37308fb8f32ca9d79d11d15.png) (или
(или ![\frac{f[i]}{f_\min}](/sites/default/files/tex_cache/d7aba164e01edf45acb87ab1365cdf0d.png) ) целевой функции по популяции.
) целевой функции по популяции.
Эту проблему решает вторая (линейная) стратегия, где срок жизни определяется исходя из значения ЦФ данной особи относительно максимального значения в популяции . Но этот метод тоже имеет свои недостатки - если в популяции много особей имеют значение ЦФ стремящееся к максимальному (![(f[i]\to f_\max)](/sites/default/files/tex_cache/b2233d20f3bca22d026ddc26eede80e1.png) ) значению, то такой подход приведет к чрезмерному увеличению размера популяции.
) значению, то такой подход приведет к чрезмерному увеличению размера популяции.
В третьей стратегии (билинейной) предпринята попытка найти компромисс между первыми двумя методами. В ней учитываются разница между сроками жизни, близкими к лучшей особи, используя информацию о среднем значении популяции. Однако в тоже время принимается во внимание минимальное и максимальное значение по популяции.