|
Раньше это можно было зделать просто нажав на тест и посмотреть результаты а сейчас никак |
Московский государственный университет путей сообщения
Опубликован: 01.06.2007 | Доступ: свободный | Студентов: 1914 / 103 | Оценка: 4.38 / 3.75 | Длительность: 22:59:00
ISBN: 978-5-9556-0094-9
Специальности: Программист
Теги:
Лекция 2:
Основы нейросетевых технологий
Итак, рассмотрим более подробно процесс локализации максимальной величины возбуждения на выходном слое, заключающейся в выделении того нейрона некоторой малой области, величина возбуждения которого максимальна. Он основан на подавлении тех сигналов возбуждения нейронов, которые не соответствуют нейрону с максимальным возбуждением. Т.е. если необходимо сконцентрировать сигнал и выделить нейрон с максимальной величиной возбуждения, это достигается с помощью подавляющих связей, с которыми действуют друг на друга "соседние" нейроны выходного слоя.
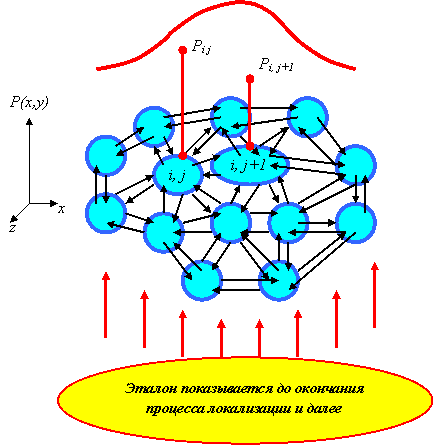
Пусть в целом над нейронами выходного слоя, условно расположенного на плоскости (x, y), можно построить непрерывную функцию их возбуждения P(x, y) (рис. 2.18), обусловленную прохождением сигналов возбуждений в сети на основе предъявленного эталона. Будем считать, что эта функция имеет один или более максимумов. Пусть Pij - значение величины возбуждения нейрона с координатами (i, j).
Каждый нейрон (i, j), действуя в своей окрестности, рассылает соседним нейронам, на их дендриты с отрицательными, не обязательно регулируемыми, весами, свою величину возбуждения Pij, первоначально полученную из сети.
Представим взаимодействие двух "близких" нейронов, например (i, j) и (i, j+1), получивших первоначально разные значения величин возбуждения из сети. Пусть Pij > Pi,j+1 . Тогда в очередном такте времени на входе нейрона (i, j) появится подавляющий сигнал Pi,j+1 , а на входе нейрона (i, j+1), и так имеющего меньшее значение величины возбуждения, - подавляющий, больший сигнал Pij .
При подтверждаемом на входном слое эталоне, т.е. при существовании на некотором отрезке времени характера и величины возбуждений, обусловливающих возбуждение выходного слоя, на выходном слое, в частности между нейронами (i, j) и (i, j+1) , происходит перераспределение величины возбуждения.
А именно: несомненно, уменьшится значение Pij , но в еще большей степени уменьшится значение Pi,j+1 . В следующем такте более "сильный" нейрон еще более "ослабит" более "слабый" нейрон, который, в свою очередь, сможет еще в меньшей степени "ослабить" более "сильный" нейрон и т.д. Более того, "слабый" нейрон может "слабеть" до тех пор, пока взвешенная сумма подаваемых ему сигналов не станет меньше его порога.
Рассматривая этот процесс в рамках взаимодействия всех нейронов области выходного слоя, можно сделать вывод о постепенной концентрации высокого уровня возбуждения, присущего одному или нескольким нейронам и определяющего один или несколько локальных максимумов.
При таком взаимодействии нейронов области возбуждения выходного слоя происходит лишь усиление сигнала наиболее возбужденного нейрона. Полное подавление возможно лишь на границе этой области. Если где-то внутри области возбуждение некоторого нейрона окажется подавленным полностью (сигнал не преодолевает порога ), то в следующем такте этот нейрон не сможет подавить сигнал того нейрона, который до того имел более слабый сигнал возбуждения. Тогда возможно появление в такой области возбуждения нового локального максимума. Таким образом, веса отрицательных связей должны способствовать максимальному усилению возбуждения того нейрона, который первоначально продемонстрировал максимальное возбуждение, при затухании возбуждения нейронов в сторону периферии.
На человеческом языке это означает: "Любое угадывание не исключает сомнений".
Однако всегда ли необходимо на выходном слое локализовать величину возбуждения?
По-видимому, такое усиление уровня возбуждения необходимо для того, чтобы единственный нейрон выходного слоя преодолел некоторую "планку" (изменяющуюся?), чтобы объявить себя ответственным за сделанный вывод или решение.
Однако следует отметить, что локализация и максимизация возбуждения на выходном слое особенно важны тогда, когда действительно необходима высокая степень определенности. Это важно в том случае, если получаемый вывод (решение) немедленно участвует в цепочке последующих, использующих его выводов.
В конце концов, все обусловлено назначением сети, решаемой задачей. Можно представить себе возможный аттракцион - реакцию фантастического чудовища на изображение, как это представлено на рис. 2.10 и рис. 2.11. По виду изображения инициируются те или иные программы действий: радости, гнева, поднятия лап, виляния хвостом и т.д. Возбуждение определенных нейронов выходного слоя связывается с запуском соответствующих программ. Величина возбуждения может являться основным параметром для этих программ. Программы не исключают друг друга, и в одном такте могут запускаться несколько программ.
Можно представить радостную модель гурмана-дегустатора (рис. 2.20), по аромату блюда определяющего состав использованных ингредиентов и приходящего в восторг или в уныние от представленного букета.
В большинстве частных задач, где нейросеть обучается с помощью "учителя", т.е. на основе действий извне при ее настройке, присутствует элемент принудительного закрепления нейронов выходного слоя за выводами. В процессе последующего обучения преимущественно с помощью весов синапсических связей добиваются адекватной реакции сети.